







![[众诚云网科技]](/uploads/allimg/20190305/c4b08346cbe8b0efae6b132139c2d72a.png)


新闻中心 
青海省民和县喇家遗址出土家养黄牛的DNA初步研究
 2023-03-13
2023-03-13 浏览次数:87次
浏览次数:87次 返回列表
返回列表来源:赵欣 吕鹏 等 社科院考古所中国考古网 2023-02-27 17:09 发表于北京
摘要:运用古DNA技术对青海省民和县喇家遗址出土11例黄牛遗存进行DNA提取和线粒体DNA控制区分析,其中属于齐家文化时期的有10例,汉代1例。全部样本都获得了所需的DNA序列,都属于家养普通牛(Bos taurus)。在这11个样本中共检测出5个单倍型,可以归属于3个不同的单倍型类群T2、T3和T4。年代为齐家文化时期的家养普通牛样本(10例)归属于2个不同的单倍型类群T3和T4,以T3为主(80.0%),T4较少(20.0%)。年代为汉代的样本(1例)属于单倍型类群T2。喇家遗址齐家文化时期家养普通牛的母系遗传结构与同时期甘青地区其他考古遗址出土家养普通牛非常相似,可能具有很近的母系来源。
喇家遗址位于青海省民和回族土族自治县官亭镇喇家村,地处黄河上游的官亭盆地,坐落在黄河河谷地带北岸的二级阶地上,地理坐标为北纬35°42′~37°09′,东经101°05′~103°01′。喇家遗址的发掘开始于1999年,中国社会科学院考古研究所、青海省文物考古研究所联合对该遗址进行多年考古发掘工作;2015年青海省文物考古研究所联合四川大学考古系、喇家遗址博物馆等单位又对喇家国家考古遗址公园即将施工区域进行了发掘。该遗址一系列重要考古发现引起了国内外学者的广泛关注,例如目前世界上最早的“面条”[1];房址内数量不等、姿态各异的人骨[2],以及他们之间的亲缘关系[3];发现了陶器、石器、骨器、玉器等重要遗物[4];尤其是喇家遗址古代环境与灾难遗存的发现[5],揭示了距今4000年前后在黄河上游地区发生的灾变事件。喇家遗址是一处以齐家文化为主的中心聚落,被评为2001年度全国十大考古新发现。碳十四测年分析显示,喇家遗址齐家文化的年代大致延续了300多年(约为公元前2300~前1900年),而该遗址中的灾难性事件发生的年代为公元前1900年前后[6]。
本文以青海省民和县喇家遗址出土古代黄牛遗存为研究对象,运用古DNA的研究方法,分析该地区古代黄牛的遗传结构特点,并与周边甘青地区以及中国其他地区古代黄牛进行比较。本文研究期望为今后甘青地区大样本的古代黄牛DNA研究提供新的材料。
一、材料与方法
(一)样本采集与保存状况初步评估
在动物考古学家进行种属鉴定等形态学研究的基础上,本文对喇家遗址发掘出土的11例属于黄牛的骨骼或牙齿遗存进行了采集,从形态上看采集部位均保存较好,利于DNA的提取和后续实验、分析工作,其保存状况可见图一。其中有10例牛骨或牙齿出土的遗迹单位属于齐家文化时期,另有1例牛骨出土的遗迹单位为2015年发掘、年代属于汉代。样本的具体信息见表一。

图一 本文采集的喇家遗址家养黄牛骨骼情况
注:图A为未经实验处理的样本照片;图B为经过去除外源DNA污染处理后的样本照片。
(二)样本处理与DNA提取
采集到的骨骼或牙齿样本,依次经过钻头机械打磨表面和截面、次氯酸钠溶液(有效氯6%)浸泡和紫外照射(紫外灯置于样本以上8cm的高度,波长254nm,UVP,美国),用以去除表面尘垢和可能存在的外源DNA污染,之后使用液氮冷冻研磨机(SPEX,美国)将骨骼或牙齿打磨成粉。在此基础上取0.2~0.5g骨粉或牙粉,使用Yang等[7]提出的硅柱离心法进行DNA提取。每个样本得到约100uL的DNA样本置于-20℃保存备用。
(三)DNA扩增与测序
首先,使用引物L1269/H1346[8]扩增线粒体基因组中12S rRNA基因,获取76bp的短片段DNA,从分子水平对所选样本进行初步种属鉴定;在此基础上选择Yang等[9]、Troy等[10]和蔡大伟等[11]设计的四对套叠引物来扩增属于黄牛样本的线粒体DNA控制区的片段并进行拼接,获得黄牛线粒体DNA控制区331bp的序列(16004~16334,包括引物长度)。使用AmpliTaq GoldTM聚合酶(Thermo fisher,美国)进行PCR扩增,具体PCR扩增体系与程序设置见参考文献[12]。扩增产物使用2%琼脂糖凝胶进行电泳检测。PCR阳性产物送到测序公司直接进行Sanger测序,正反引物双向测序。基于本课题组在古代牛骨DNA的实验经验[13],在本实验的PCR扩增过程中全部未使用BSA(牛血清白蛋白)。
(四)数据分析
使用Chromas Pro软件对获得的DNA序列进行序列信号的读取和检测,使用Clustal X2软件[14]对DNA序列进行比对,使用BioEdit软件[15]进行编辑和拼接。使用MEGA 7软件[16]构建系统发育树,详见图二。使用Dna-SP(v.6.12.03)软件[17]和Arlequin(v. 3.5)软件[18]计算单倍型多样性与核苷酸多样性;使用PopART(v. 1.7)软件[19]构建中介网络图(图三A),使用TempNet R程序[20]构建甘青地区家养黄牛的分层网络图(图三B,按照遗址年代划分层次);以遗址为单位计算单倍型类群分布频率(图四),选择中国考古遗址出土家养普通牛作为对比序列,包括:山西陶寺、山西周家庄、河南花地嘴、河南望京楼、河南二里头、河南殷墟孝民屯、陕西泉护村、陕西石峁、吉林后套木嘎、内蒙古大山前、青海长宁、宁夏打石沟、宁夏沙塘北塬、宁夏王大户、宁夏中庄、宁夏九龙山、新疆小河等[21]。
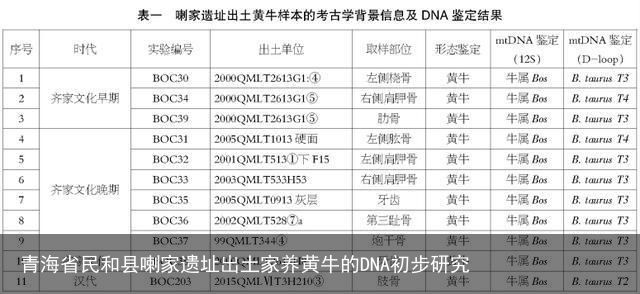
图二 系统发育邻接树
注:本文研究样本用方块表示,用BOC标注编号。对比序列来自GenBank数据库,用圆圈表示,序列编号为GenBank号,包括:普通牛单倍型类群T1(GenBank:JN817351和GenBank:MF475809)、普通牛单倍型类群T2(GenBank:KT184456和GenBank:MF475825)、普通牛单倍型类群T3(GenBank:V00654)、普通牛单倍型类群T4(GenBank:DQ124372)、瘤牛单倍型类群I(GenBank:EU177868和GenBank:EU177870)、亚洲原始牛单倍型类群C(用C表示,GenBank:MG279478)、欧洲原始牛(用P表示,GenBank:DQ915537)、牦牛(用G表示,GenBank:GQ464297)和水牛(用B表示,GenBank:NC_006295和GenBank:AY195595)。全部节点都标注自展值,比例尺表示每一个位点的核苷酸替代率。

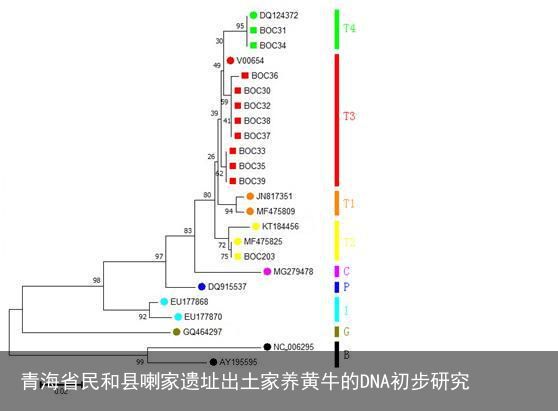
图三 中介网络图
注:每一个圆圈代表一个单倍型,圆圈的大小与共享这一单倍型的样本数量成正比。
图三A:基于中国古代家养普通牛线粒体DNA控制区序列构建的中介网络图(不同颜色代表不同遗址)。
图三B:基于中国甘青地区古代家养普通牛线粒体DNA控制区序列构建的分层网络图,按照遗址年代划分为两个层次:下层(红色)为新石器时代至青铜时代,上层(蓝色)为春秋战国时期。不同年代共享的单倍型由竖直的实线相连,圆圈中的数字表示共享这一单倍型的样本数量;Hap_1、Hap_3和Hap_18分别对应图三A中的单倍型。
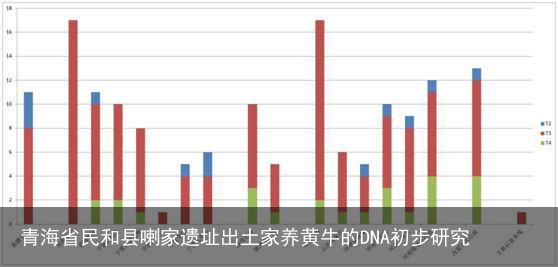
图四 中国考古遗址出土家养普通牛单倍型类群分布频率
(五)污染的防止与序列真实性验证
残存在古代生物遗骸中的遗传物质DNA分子具有含量极低、高度降解、广泛损伤等几个特点。基于此,使得古DNA研究极易受到外源DNA污染。外源DNA污染的防止与去除,样本DNA序列真实性验证等,是进行古DNA研究的前提,也是保证古DNA研究可靠性的最大问题。本文全部实验都严格按照古DNA操作规范进行。所有实验都在专门的古DNA实验室完成,且每次DNA提取与PCR扩增都设置空白对照,以判断是否在实验过程中存在外源污染。实验中所有样本都提取3次以上,保证不同次提取产物结果相同。所有的DNA片段都进行正反引物双向测序且得到完全一致的结果,用以识别和排除实验中可能存在的误差。虽然BSA(牛血清白蛋白)可以有效地提高PCR扩增成功率,在古DNA实验中应用广泛。但是BSA是从牛组织中提取的产物,本文的研究样本是古代黄牛,考虑到无法排除BSA可能会带来外源DNA污染,因此本文实验未使用BSA,从实验试剂方面排除了外源污染的可能。
二、结果
本文实验设置的全部空白对照都是阴性,未检测到实验污染;获得的DNA序列都得到了重复验证,显示出所获得的DNA数据的可信性。
在采集的11例古代样本中,全部获得了线粒体基因组中12S rRNA基因的部分DNA序列(引物L1269/H1346,去除两端引物后序列长度为76bp),在GenBank数据库中(http://www.ncbi.nlm.nih.gov)对这些DNA序列进行了BLAST共享序列搜索,结果发现全部样本的共享序列都属于牛属(Bos)。同样这11例样本都获得了线粒体DNA控制区的285bp序列(16029~16313,去除两端引物),在GenBank数据库中(http://www.ncbi.nlm.nih.gov)对这些DNA序列进行了BLAST共享序列搜索,结果发现全部样本都属于家养普通牛(Bos taurus)。
以V00654[22]作为参考序列,将这11个线粒体DNA控制区序列与之进行比对,实验结果见表二。共检测出10个多态性位点,其中有9个为转换,1个为颠换(16057G→C),没有插入或缺失,且转换在嘧啶之间的比例略高占66.7%(6/9)。在这11个序列中共检测出5个单倍型,其中BOC30、BOC32、BOC37和BOC38, BOC33、BOC35和BOC39,BOC31和BOC34分别共享有相同的单倍型。虽然BOC36在该遗址中无共享序列,但是与BOC30这一单倍型仅有一个位点的差异(16068T→C),表现出较近的母系亲缘关系。根据家养普通牛单倍型类群划分的已有研究成果[23],基于线粒体DNA控制区序列变异模式,本文发现的5个单倍型可以归属于3个不同的单倍型类群T2、T3和T4,以T3为主占72.7%,T4仅发现2个个体(BOC31和BOC34)占18.2%,T2仅发现1个个体(BOC203)占9.1%,未见其他单倍型类群,结果详见表二和图二。
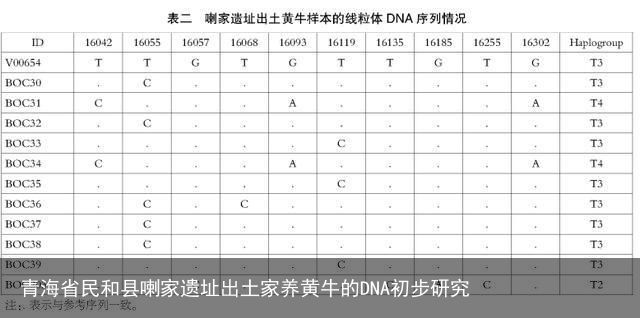
由于BOC203出土的遗迹单位属于汉代,其他样本出土的遗迹单位都属于齐家文化时期,二者年代相距较远,应该依据其年代分为两个组分别进行统计更为妥当。因此将同时期样本进行单倍型类群分布频率计算,结果如下:属于齐家文化时期的10个家养普通牛中共发现4个单倍型可以归属于2个不同的单倍型类群T3和T4,以T3为主占80.0%,T4仅发现2个个体(BOC31和BOC34)占20.0%,未见其他单倍型类群;年代为汉代的样本仅1例(BOC203)属于单倍型类群T2。
三、讨论
古DNA研究的可行性主要依赖于考古遗址出土样本DNA的保存状况。青海地处高海拔地区,气候相对低温干燥,利于古DNA的保存,是本文实验中DNA提取成功率达到100%的主要环境因素。
作为“六畜”之一,黄牛的饲养在我国有着悠久的历史。牛(Bovine)为牛亚科下的一个族,属牛族,包括牛属(Bos)、水牛属(Bubalus)等,又分为多个种,既有野生种也有驯化种。在中国通常所说的黄牛[24],主要包括普通牛(Bos taurus)和瘤牛(Bos indicus)两个种,都属于驯化种。虽然学术界对家养黄牛在中国最早出现的时间这个问题并未达成一致,但是一般认为家养普通牛大约是在公元前3600~前2000年左右从西亚地区传入中国[25]。关于瘤牛在中国最早出现的时间,并不能确定,因为目前还没有发现明确属于瘤牛的骨骼遗存材料。
本文采集的喇家遗址出土黄牛样本全部属于家养普通牛(表二、图二)。依据考古发掘背景信息可知,属于齐家文化时期的有10例,其中3例可归属到齐家文化早期(BOC30、BOC34和BOC39),6例可归属到齐家文化晚期(BOC31、BOC32、BOC33、BOC35、BOC36和BOC37),BOC38属于齐家文化时期但不能确定是早期亦或晚期,BOC203属于汉代。属于齐家文化时期的10例家养普通牛中共发现4个单倍型,归属于2个不同的单倍型类群T3和T4,以T3为主占80.0%,T4仅发现2个个体(BOC31和BOC34)占20.0%。以齐家文化早期和晚期分别统计可以看出,属于齐家文化早期的3例家养普通牛中共发现3个不同的单倍型,归属于2个不同的单倍型类群T3和T4,以T3为主占66.7%,T4发现1个个体占33.3%;属于齐家文化晚期的6例家养普通牛中共发现4个单倍型,同样可以归属于2个不同的单倍型类群T3和T4,以T3为主占83.3%,T4发现1个个体占16.7%。从所发现的单倍型(即共享序列)来看,齐家文化早期的3个样本在齐家文化晚期的家养普通牛中都找到了共享有相同序列的样本。因此,可以认为齐家文化早期与晚期家养普通牛的母系遗传结构相似、并且具有非常近的母系亲缘连续性。由于喇家遗址齐家文化早期与晚期家养普通牛的遗传结构相似,因此下文中将喇家遗址齐家文化家养普通牛作为一个群体进行统计分析。目前来看喇家遗址在齐家文化时期没有发现属于单倍型类群T2的样本,属于单倍型类群T2的样本在该遗址出现的年代较晚在汉代、且仅有1例样本(BOC203),下文在进行统计分析时将不再包括这1例汉代黄牛样本。
从图三A中可以看出,在喇家遗址发现的5个单倍型(Hap_1~Hap_5)中,有4个单倍型(Hap_1~Hap_3,Hap_5)在中国其他古代家养普通牛中都有发现。如前文所述Hap_4(本文实验编号BOC36)虽然在其他中国古代家养黄牛中未找到共享序列,但是与Hap_1这一单倍型仅有一个位点的差异(16068T→C),表现出比较近的母系亲缘关系。Hap_5(本文实验编号BOC203)属于汉代,将在下文中单独讨论。在其他的3个单倍型(Hap_1~Hap_3)中,Hap_1和Hap_3属于家养普通牛T3中最常见也是所占比例非常高的单倍型,在中国多个考古遗址的家养普通牛中都有发现,特别是在新石器时代晚期至青铜时代早期甘青地区的几个遗址中都有发现。Hap_2属于家养普通牛T4,在大山前、望京楼和殷墟孝民屯三个遗址中有发现,但是在甘青地区其他遗址中未见到。
将中国古代家养黄牛以遗址为单位进行单倍型类群统计(图三A、图四)发现,与喇家遗址齐家文化家养普通牛母系遗传结构特征相似(同时存T3和T4、且以T3为主)的遗址主要包括山西陶寺(龙山时期)、山西周家庄(龙山时期)、陕西泉护村(青铜时代)、陕西石峁(龙山末期至二里头时期)、宁夏打石沟(新石器时代)和宁夏沙塘北塬(新石器时代晚期至青铜时代早期)。一方面,从年代来看,在这些遗址中,除陕西泉护村属于青铜时代外,其他几个遗址的年代都可以早到新石器时代晚期,说明在这一时期中国北方地区的古代家养黄牛的母系遗传结构比较简单、且各遗址比较相似。除上述遗址外年代可以早到新石器时代的遗址仅有吉林后套木嘎遗址,但是该遗址仅发现1例家养普通牛属于T3并不具备统计学意义。另一方面,从地域来看,位于甘青地区且年代可以早到新石器时代至青铜时代早期的遗址共有4个,为青海长宁、青海喇家、宁夏打石沟和宁夏沙塘北塬,除长宁遗址家养普通牛的遗传结构更为简单(仅存在T3)外,喇家、打石沟和沙塘北塬三个遗址出土的家养普通牛的母系遗传结构非常相似,并且在这四个遗址中都存在共享有相同单倍型的现象(Hap_1和Hap_3)。长宁遗址位于青海省西宁市大通县长宁乡长宁村西南约3千米处,湟水流域一处重要的以齐家文化(距今4200~3800年左右)为主的大型聚落遗址,处于新石器时代向青铜时代的过渡阶段[26]。打石沟遗址位于宁夏彭阳县古城镇店洼村水库东50米的坡地上,发掘者研究认为该遗址的文化特征属于新石器时代晚期客省庄二期文化(距今4300~4000年左右)[27]。沙塘北塬遗址所处地理位置与打石沟较近,最新研究认为应该属于常山下层文化向齐家文化的一个过渡类型(公元前2200~前1900年之间)[28]。喇家遗址主体属于齐家文化,距离上述三个遗址位置不远,从考古学文化区系上讲都属于甘青地区,年代亦与上述三个遗址相近。虽然遗址所属考古学文化有异,但是新石器时代至青铜时代早期在甘青地区考古遗址出土的家养普通牛具有非常近的母系亲缘关系,不排除有很近的母系来源。
进入春秋战国时期以后,属于甘青地区的有宁夏王大户、中庄和九龙山共三个遗址黄牛DNA研究成果已经发表。它们仍然都是家养普通牛,但是与甘青地区早期家养黄牛相比,其母系遗传结构有比较明显的变化(图三A、图四)。从单倍型类群来看,除王大户遗址仅发现1例家养普通牛属于T3不具备统计学意义以外,中庄和九龙山都出现了新的单倍型类群T2,并且直到汉代依然存在(喇家);而原来一直存在的单倍型类群T4在春秋战国时期的考古遗址中并未发现。该单倍型类群分布频率仅与新疆小河遗址相似,与已发表的其他中国考古遗址都不同。
除了有单倍型类群分布频率的较大变化外,就单倍型类群T3内部而言,从新石器时代到春秋战国时期也有较大改变(图三A、图三B)。在新石器时代至青铜时代早期最主要的单倍型是Hap_1和Hap_3,分别占到甘青地区这一时期全部黄牛的33.3%(15/45)和42.2%(19/45),分别占到单倍型类群T3的37.5%(15/40)和47.5%(19/40)。到了春秋战国时期后,未发现有单倍型Hap_1,属于单倍型Hap_3的也只有1个个体(中庄遗址),在甘青地区单倍型Hap_3是唯一一个新石器时代至青铜时代早期和春秋战国时期同时存在的单倍型;而新石器时代至青铜时代早期并未发现的单倍型Hap_18在王大户、中庄和九龙山这三个遗址中都有发现,占到甘青地区春秋战国时期全部黄牛的33.3%(4/12),占到T3的44.4%(4/9)。由此可见进入青铜时代以后,特别是到了春秋战国时期以后,甘青地区家养黄牛的母系遗传结构发生了比较大的改变(图三B),这种改变已经超过了中国其他地区的考古遗址。在其他地区考古遗址出土家养普通牛中,单倍型Hap_1和Hap_3比较常见,特别是在中原地区,从新石器时代到青铜时代都占有较高的比例;并且随着时间的发展,进入青铜时代以后,多数遗址发现了新的单倍型类群T2,但是单倍型类群T4也依然存在。由此推测不同的地区对于家养黄牛饲养品种具有不同的选择性。进入青铜时代以后甘青地区家养黄牛多样性的变化,不排除是从西亚、中亚等地区不断地引进新品种的结果。
属于单倍型类群T2的样本(BOC203)在喇家遗址出现的年代较晚在汉代、且仅有1例,不具备统计学分析的意义。但是从图三A中可以看出,与其共享有相同单倍型(Hap_5)的样本还有2例,分别发现于中庄和九龙山,都为甘青地区春秋战国时期的考古遗址。或许可以说从春秋战国时期到汉代甘青地区古代黄牛的母系遗传具有一定的连续性。
目前为止还没有证据能够证明龙山时代晚期是家养黄牛在中国出现的最早时期,但是动物考古学与古DNA研究都可以证明在龙山时代晚期已经饲养黄牛,并且这一时期中国北方考古遗址出土的家养黄牛遗传结构相对简单。同时有的遗址中又伴出有野生原始牛和圣水牛[29],推测当时黄牛的饲养数量和饲养能力相对较低,可能处于黄牛饲养的早期阶段[30]。但是到了公元前2000年前后单倍型类群T2、T3和T4在中国已全部存在[31],并且这种遗传模式在中原地区延续下来。
综合已发表的古DNA研究成果认为,中国北方地区青铜时代黄牛均为普通牛,未见瘤牛;中国古代家养黄牛对现代东亚家养普通牛具有重要的遗传贡献。中国家养黄牛最早由近东地区引入中国,近东起源的普通牛可能经过两条路线进入中国。经由河西走廊、最终到达中原地区的“西线”,以及经欧亚草原通道到达中国北方的东部地区的“北线”[32]都有可能是黄牛在中国扩散的主要路线,并且暗示着这两条路线的扩散活动可能是同时进行的[33]。本文关于喇家遗址古代黄牛的DNA研究结果,为家养普通牛通过“西线”传播进入中国又提供了新的证据。与“西线”相比,目前来看“北线”及相关区域的材料太少,这将会是今后工作的重点。
中国甘青地区处于北方农牧交错带及河西走廊沿线,考古学研究表明,自古以来就是东西文化交流的要冲和活跃地带,是重要的生业技术传播的交通要道。依据各类家养动物在甘青地区出现的时间、顺序和特征,可以看出来新石器时代的甘青地区是一个接纳来自不同方向的多种家养动物的区域[34]。本课题组在对喇家遗址出土汉代家马的线粒体DNA遗传研究中推测,作为丝绸之路的一部分,喇家所在的甘青地区在沟通中原地区和新疆地区中处于独特的地理位置,是家马从新疆传入中原的必经之路。结合本文研究结果可知,对于探索早期家养动物的起源与驯化,揭示从新石器时代向青铜时代过渡时期黄河上游地区不同考古学文化对这些家养动物的饲养与使用策略的多样化,甘青地区具有至关重要的作用。
本文从线粒体DNA母系遗传的角度着手对喇家遗址出土黄牛进行古DNA研究,成功率达到了100%,显示出这批材料保存状况良好,喇家遗址的埋藏环境适于古代动物遗存及其DNA的保存。下一步我们将对这批材料开展以下一代测序技术为核心的基因组学研究,获取古代黄牛基因组数据,并抓出其中有用的遗传信息,运用基因组学大数据为解决考古学问题提供更多的线索,以弥补单一从母系遗传角度进行分析而有可能忽略父系方面遗传信息的问题。
四、 结论
考古学家研究认为被誉为东方的庞贝古城的喇家遗址处于半农半牧的经济形态[35],该遗址大量动物遗存的发现,为研究甘青地区早期家养动物的饲养与传播提供重要的动物考古学信息。本文对喇家遗址出土黄牛的DNA研究,虽样本量不大,且主要是通过线粒体DNA母系遗传结构进行分析,但仍得出以下认识:
(1)喇家遗址出土黄牛样本全部属于家养普通牛。属于齐家文化时期的家养普通牛可以归属于线粒体DNA单倍型类群T3和T4,以T3为主,T4次之;齐家文化早期与晚期家养普通牛的母系遗传结构相似、并且具有非常近的母系亲缘连续性。另有1例样本年代为汉代,属于单倍型类群T2。
(2)新石器时代至青铜时代早期甘青地区考古遗址出土的家养普通牛具有非常近的母系亲缘关系,不排除有很近的母系来源。但是春秋战国时期该地区几个考古遗址出土家养普通牛的母系遗传结构有了比较明显的变化。不排除青铜时代以后甘青地区家养普通牛的多样性变化,是从西亚、中亚等地区不断地引进新品种的结果。
致谢:感谢中国社会科学院考古研究所、青海省文物考古研究所、四川大学考古系、喇家遗址博物馆等为本研究提供了宝贵的样本;感谢编辑部老师提出的宝贵意见。
向上滑动阅读正文[1]. a. Lü H, Yang X, Ye M, et al. Culinary archaeology: Millet noodles in Late Neolithic China. Nature, 2005, 437: 967-tu;b. 吕厚远、李玉梅、张健平等:《青海喇家遗址出土4000年前面条的成分分析与复制》,《科学通报》2015年第60卷第8期,第744~756页。[2]. a. 中国社会科学院考古研究所甘青工作队、青海省文物考古研究所:《青海民和县喇家遗址2000年发掘简报》,《考古》2002年第12期,第12~25页;b. 中国社会科学院考古研究所、青海省文物考古研究所:《青海民和喇家史前遗址的发掘》,《考古》2002年第7期,第3~5页。[3]. a. 王明辉:《青海民和喇家遗址人骨及相关问题》,《考古》2002年第12期,第26~28页;b. 赵春燕、王明辉、叶茂林:《青海喇家遗址人类遗骸的锶同位素比值分析》,《人类学学报》2016年第35卷第2期,第212~222页;c. Gao S, Yang Y, Xu Y, et al. Tracing the genetic history of the Chinese people: mitochondrial DNA analysis of a Neolithic population from the Lajia Site. American Journal of Physical Anthropology, 2007, 133: 1128-1136;d. 张雪莲、叶茂林:《喇家遗址先民食物的初步探讨——喇家遗址灾难现场出土人骨的碳氮稳定同位素分析》,《南方文物》2016年第4期,第197~202页。[4]. a. 叶茂林:《青海民和县喇家遗址出土齐家文化玉器》,《考古》2002年第12期,第89~90页;b. 青海省文物考古研究所等:《青海民和喇家遗址2015年发掘收获》,见国家文物局主编:《2015中国重要考古发现》,北京:文物出版社,2016年4月第1版,第40~44页。[5]. 夏正楷、杨晓燕、叶茂林:《青海喇家遗址史前灾难事件》,《科学通报》2003年第48卷第11期,第1200~1204页。[6]. 张雪莲、叶茂林、仇士华等:《民和喇家遗址碳十四测年及初步分析》,《考古》2014年第11期,第91~104页。[7]. Yang D Y, Eng B, Waye J S. Technical Note: Improved DNA extraction from ancient bones using silica-based spin columns. American Journal of Physical Anthropology, 1998, 105(4): 539-543.[8]. Rollo F, Ubaldi M, Ermini L, et al. Ötzi’s last meals: DNA analysis of the intestinal content of the Neolithic glacier mummy from the Alps. PNAS, 2002, 99(20): 12594-12599. doi/10.1073/pnas.192184599.[9]. Yang D, Liu L, Chen X, et al. Wild or domesticated: DNA analysis of ancient water buffalo remains from north China. Journal of Archaeological Science, 2008, 35(10): 2778-2785. http://dx.doi.org/10.1016/j.jas.2008.05.010.[10]. Troy C S, MacHugh D E, Bailey J F, et al. Genetic evidence for Near-Eastern origins of European cattle. Nature, 2001, 410(6832): 1088-1091. http://dx.doi.org/10.1038/35074088.[11]. Cai D, Sun Y, Tang Z, et al. The origins of Chinese domestic cattle as revealed by ancient DNA analysis. Journal of Archaeological Science, 2014, 41: 423-434. http://dx.doi.org/10.1016/j.jas.2013.09.003.[12]. 赵欣、顾万发、吴倩等:《河南省郑州地区青铜时代遗址出土牛骨的DNA研究》,《南方文物》2018年第4期,第126~134+40页。[13]. a. 赵欣、顾万发、吴倩等:《河南省郑州地区青铜时代遗址出土牛骨的DNA研究》,《南方文物》2018年第4期,第126~134+40页;b. 赵欣、李志鹏、东晓玲等:《河南安阳殷墟孝民屯遗址出土家养黄牛的DNA研究》,《第四纪研究》2020年第40卷第2期,第321~330页;c. 赵欣、东晓玲 、刘铭等:《宁夏隆德县沙塘北塬遗址出土家养黄牛的DNA研究》,《南方文物》2021年第5期,第167~172页。[14]. Larkin M A, Blackshields G, Brown N P, et al. Clustal W and clustal X version 2.0. Bioinformatics, 2007, 23: 2947-2948. http://dx.doi.org/10.1093/bioinformatics/btm404.[15]. Simmons M P. Independence of alignment and tree search. Molecular Phylogenetics and Evolution, 2004, 31(3): 874-879.[16]. Kumar S, Stecher G, Tamura K. MEGA 7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Molecular Biology and Evolution, 2016, 33(7): 1870-1874.[17]. Librado P, Rozas J. DnaSP v5: a software for comprehensive analysis of DNA polymorphism data. Bioinformatics, 2009, 25: 1451-1452.[18]. Excoffier L, Lischer H E L. Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Molecular Ecology Resources, 2010, 10: 564-567.[19]. Bandelt H-J, Forster P, Röhl A. Median-joining networks for inferring intraspecific phylogenies. Molecular Biology and Evolution, 1999, 16(1): 37-48.[20]. Prost S, Anderson C N. TempNet: a method to display statistical parsimony networks for heterochronous DNA sequence data. Methods in Ecology and Evolution, 2011, 2(6): 663-667.[21]. a. Brunson K, Zhao X, He N, et al. New insights into the origins of oracle bone divination: Ancient DNA from Late Neolithic Chinese bovines. Journal of Archaeological Science, 2016, 74: 35-44. http://dx.doi.org/10.1016/j.jas.2016.08.008;b. Cai D, Sun Y, Tang Z, et al. The origins of Chinese domestic cattle as revealed by ancient DNA analysis. Journal of Archaeological Science, 2014, 41: 423-434. http://dx.doi.org/10.1016/j.jas.2013.09.003.c. 赵欣、顾万发、吴倩等:《河南省郑州地区青铜时代遗址出土牛骨的DNA研究》,《南方文物》2018年第4期,第126~134+40页;d. 赵欣、李志鹏、东晓玲等:《河南安阳殷墟孝民屯遗址出土家养黄牛的DNA研究》,《第四纪研究》2020年第40卷第2期,第321~330页;e. 蔡大伟、胡松梅、孙洋等:《陕西泉护村古代黄牛的分子考古学研究》,《考古与文物》2014年第5期,第116~120页;f. 蔡大伟、胡松梅、孙玮璐等:《陕西石峁遗址后阳湾地点出土黄牛的古DNA分析》,《考古与文物》2016年第4期,第122~127页;g. Cai D, Zhang N, Zhu S, et al. Ancient DNA reveals evidence of abundant aurochs (Bos primigenius) in Neolithic Northeast China. Journal of Archaeological Science, 2018, 98: 72-80;h. Cai D, Luan Y, Gao Y, et al. Molecular archaeological research on ancient cattle from the early bronze age changning site, Qinghai Province. Asian archaeology, 2015, 3: 167-175;i. 蔡大伟、张乃凡、朱存世等:《宁夏新石器时代晚期至春秋战国时期黄牛的分子考古学研究》,《边疆考古研究》(第23辑),北京:科学出版社,2018年,第315~329页;j. 赵欣、东晓玲 、刘铭等:《宁夏隆德县沙塘北塬遗址出土家养黄牛的DNA研究》,《南方文物》2021年第5期,第167~172页;[22]. Anderson S, de Bruijn M H L, Coulson A R, et al. Complete sequence of bovine mitochondrial DNA conserved features of the mammalian mitochondrial genome. Journal of Molecular Biology, 1982, 156(4): 683-717. http://dx.doi.org/10.1016/0022-2836(82)90137-1.[23]. a. Anderson S, de Bruijn M H L, Coulson A R, et al. Complete sequence of bovine mitochondrial DNA conserved features of the mammalian mitochondrial genome. Journal of Molecular Biology, 1982, 156(4): 683-717. http://dx.doi.org/10.1016/0022-2836(82)90137-1.b. Mannen H, Tsuji S, Loftus R T, et al. Mitochondrial DNA variation and evolution of Japanese black cattle (Bos taurus). Genetics, 1998, 150(3): 1169-1175;c. Mannen H, Kohno M, Nagata Y, et al. Independent mitochondrial origin and historical genetic differentiation in north eastern Asian cattle. Molecular Phylogenetics and Evolution, 2004, 32(2): 539-544. http://dx.doi.org/10.1016/j.ympev.2004.01.010;d. Troy C S, MacHugh D E, Bailey J F, et al. Genetic evidence for Near-Eastern origins of European cattle. Nature, 2001, 410(6832): 1088-1091. http://dx.doi.org/10.1038/35074088.[24]. a. 郭郛、李约瑟、成庆泰:《中国古代动物学史》,北京:科学出版社,1999年;b. 邱怀主编:《中国黄牛》,北京:农业出版社,1992年。[25]. a. Flad R K, Yuan J, Li S. Zooarchaeological evidence for animal domestication in northwest China. In: Madsen D B, Chen F H, Gao X. (eds.) Late Quaternary Climate Change and Human Adaptation in Arid China. Amsterdam; London: Elsevier. 2007: pp. 167-203. https://doi.org/10.1016/S1571-0866(07)09012-4;b. 吕鹏:《论中国家养黄牛的起源》,《动物考古(第一辑)》,北京:文物出版社,2010年,第152~176页;c. Lu P, Brunson K, Yuan J, et al. Zooarchaeological and genetic evidence for the origins of domestic cattle in ancient China. Asian Perspectives, 2017, 56(1): 92-120.[26]. 任晓燕:《青海长宁遗址抢救性考古取得重要成果》,《中国文物报》第2版,2006-12-20。[27]. 马强、郭家龙:《彭阳县打石沟新石器时代遗址》,见中国考古学会编:《中国考古学年鉴(2014)》,北京:中国社会科学院出版社,2015年,第464页。[28]. 宁夏文物考古研究所:《宁夏隆德县沙塘北塬遗址2016年发掘简报》,《考古》2020年第4期,第3~21页。[29]. a. Brunson K, Zhao X, He N, et al. New insights into the origins of oracle bone divination: Ancient DNA from Late Neolithic Chinese bovines. Journal of Archaeological Science, 2016, 74: 35-44. http://dx.doi.org/10.1016/j.jas.2016.08.008.b. Cai D, Zhang N, Zhu S, et al. Ancient DNA reveals evidence of abundant aurochs (Bos primigenius) in Neolithic Northeast China. Journal of Archaeological Science, 2018, 98: 72-80. [30]. 赵欣:《中国考古遗址出土黄牛骨骼的分子考古学研究初探》,《科技考古(第六辑)》,北京:科学出版社,2021年,第138~151页。[31]. Cai D, Sun Y, Tang Z, et al. The origins of Chinese domestic cattle as revealed by ancient DNA analysis. Journal of Archaeological Science, 2014, 41: 423-434. http://dx.doi.org/10.1016/j.jas.2013.09.003.[32]. 赵志军:《小麦东传与欧亚草原通道》,《三代考古(三)》,北京:科学出版社,2009年,第456~459页。[33]. 蔡大伟、张乃凡、朱存世等:《宁夏新石器时代晚期至春秋战国时期黄牛的分子考古学研究》,《边疆考古研究》(第23辑),北京:科学出版社,2018年,第315~329页。[34]. 傅罗文、袁靖、李水城:《论中国甘青地区新石器时代家养动物的来源及特征》,《考古》2009年第5期,第80~86+113页。[35]. 叶茂林:《齐家文化农业发展的生态化适应:原始草作农业初探——以青海喇家遗址为例》,《农业考古》2015年第6期,第19~26页。
*作者:赵欣 中国社会科学院考古研究所科技考古中心、西蒙菲莎大学(Simon Fraser University)考古学系古DNA实验室,吕鹏 东晓玲 中国社会科学院考古研究所科技考古中心,张桦 西蒙菲莎大学(Simon Fraser University)考古学系古DNA实验室,叶茂林 中国社会科学院考古研究所,蔡林海 杜玮 马骞 甄强 青海省文物考古研究所,袁靖 中国社会科学院考古研究所科技考古中心、复旦大学科技考古研究院,杨东亚 中国社会科学院考古研究所科技考古中心、西蒙菲莎大学(Simon Fraser University)考古学系古DNA实验室;原文刊于《南方文物》2022年第6期。
责编:韩翰 荼荼
